本網站已搬家!
自 2014 年 3 月起,我們將網站遷移到新伺服器:http://web.ntnu.edu.tw/~treehopper/。本網頁內容已不再更新。請更新您的瀏覽器書籤亦或任何相關網址。

系統分類學期刊 2004 vol.53 isssue 3 封面報導本實驗室研究的角蟬(與其它昆蟲) (Lin et al 2004).
研究主題
目錄
研究興趣
1. 角蟬生物學

2000 年在巴拿馬 Chiriqui Grande 拍攝的 Membracis 角蟬雌蟲 (photo by CPL).
背覆奇異盔甲的迷你怪獸—角蟬是個龐大的昆蟲家族,牠們彷彿是大自然開的玩笑,有著奇特古怪的外型與生活史,更妙的是牠與螞蟻之間的互動,堪稱是共演化的最佳見證 (角蟬與螞蟻的共演化) 。 角蟬是一群半翅目植食性昆蟲, 具有穿刺性口器, 以吸取植物篩管和導管的汁液為食. 大多數的角蟬可用胸部背上奇特且增大的構造 (前胸背板) 輕易和相似的昆蟲區分, 例如葉蟬,浮塵子,和泡沫蟬. 和許多半翅目昆蟲一樣, 角蟬具有濾網式的中腸, 可以將樹液中的營養物質 (胺基酸和碳水化合物) 萃取出來, 含糖多餘的水份以蜜露的型式排出, 這些蜜露吸引許多膜翅目社會型昆蟲如螞蟻,蜜蜂,和黃蜂來覓食. 由於分泌蜜露, 很多角蟬和螞蟻有互利共生關係而受到螞蟻的保護.(讀更多有關角蟬的生物學, Wood 1993, Lin 2006)
2. 振動溝通系統
藉由介質傳遞的振動訊號是群居性角蟬在寄主植物上生存不可或缺的工具. 有些角蟬稚蟲在捕食者接近時會向母蟲發出求救訊號. 而有些角蟬在發現覓食處後會向兄弟姐妹發出訊號. 另外很多種角蟬則發出許多功能未知的振動訊息. 與密蘇里大學生物系 Reginald Cocroft 合作, 在國家地理雜誌社探索研究計畫贊助下, 我們發現角蟬稚蟲有非常豐富的振動溝通系統 . (參見在美國國家公共電台播放的 '亞馬遜河邊緣之旅' , NPR Morning Edition, May 24, 2006). 這些新發現提供了探索振動溝通系統如何和角蟬的社會行為和生態同步演化獨特的新視野.
3. 系統分類與親緣關係
“ 物種親緣關係比較像一個統計分佈, 由眾多親緣樹 (基因樹) 組成, 每一個顯示不同的演化關係. ”
—Wayne Maddison, 1995
i. 角蟬亞科的親緣樹

角蟬亞科中的角蟬種類代表 (from Lin et al 2004).
系統分類學的一個主要的研究課題是探討物種之間的關係和演化歷史.我們以數個粒線體和核基因DNA序列重建新熱帶角蟬亞科(包含5族, 38屬, 約447已命名種)的親緣關係. 演化樹重現了各族和屬之間的親緣遠近, 且演化樹中各類群演化分岐處有很高的支持度. (Lin et al 2004).
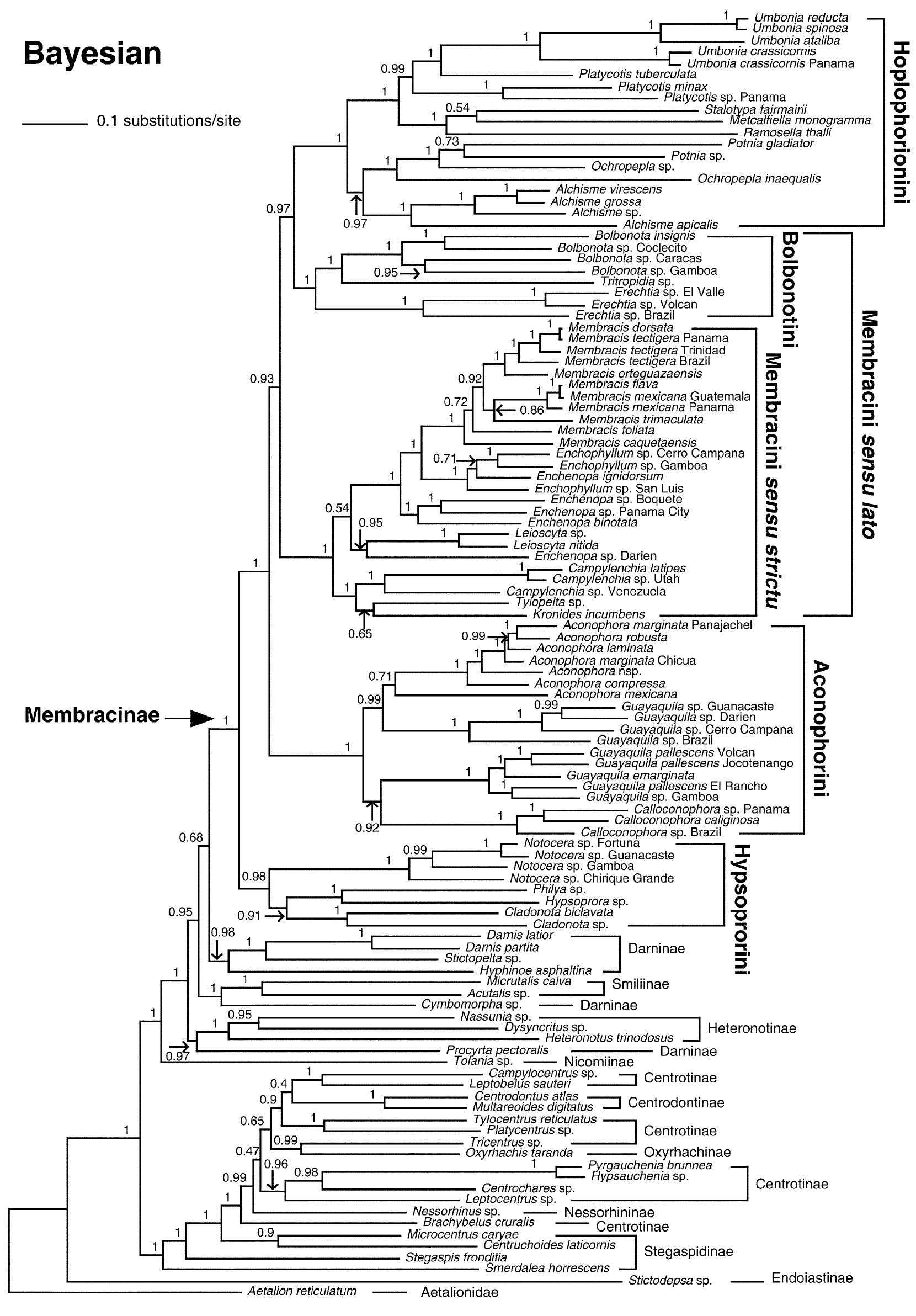
由分子特徵重建的角蟬亞科演化樹 (Lin et al 2004).
ii. Enchenopa binotata 角蟬種群親緣樹
撰寫中...
iii. 粒線體基因和核基因在親緣分析中的效能
撰寫中...
iv. 臺灣深山屬鍬形蟲親緣關係
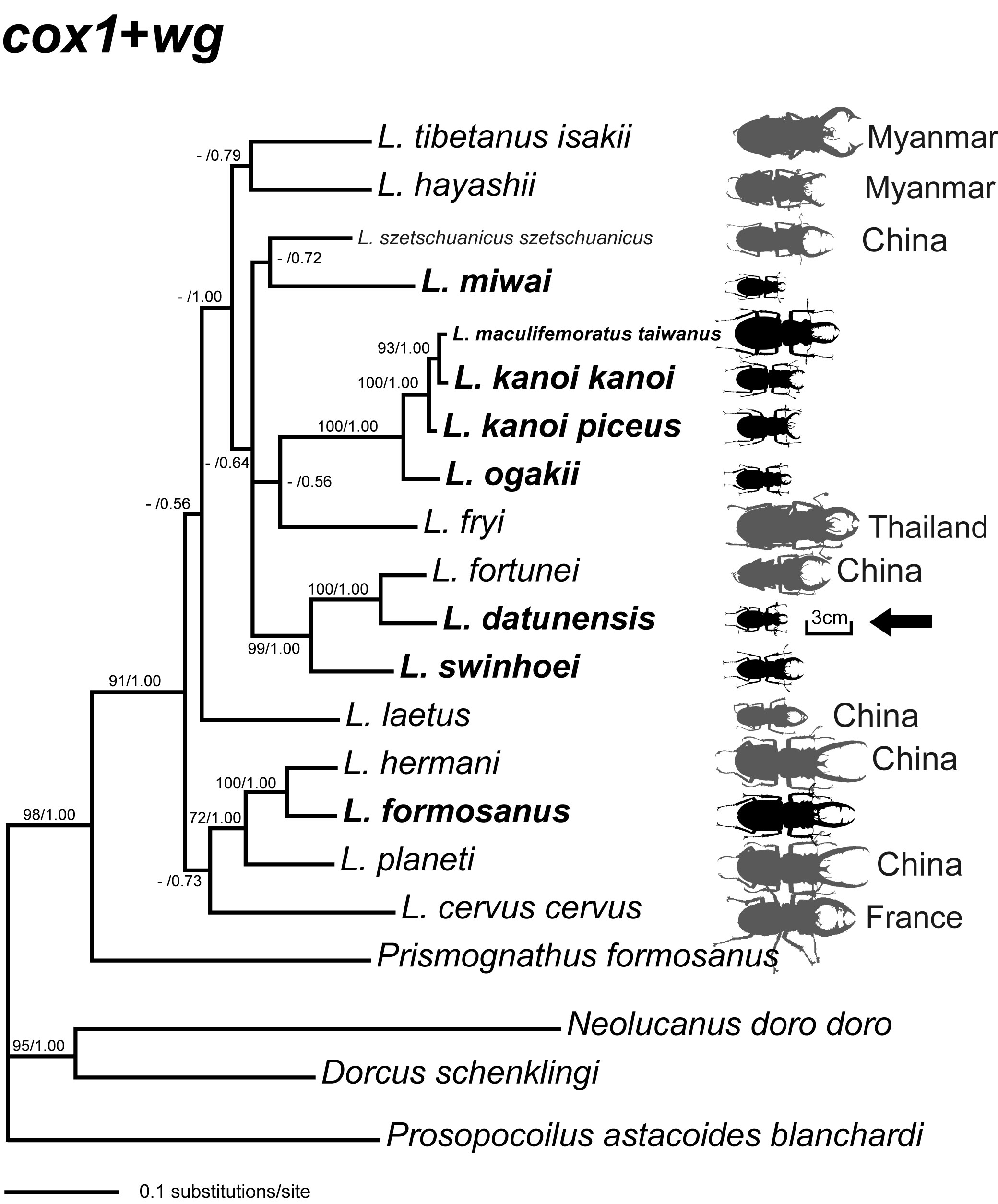
臺灣深山屬 Lucanus 鍬形蟲演化樹 (from Lin et al 2009).
大屯姬深山鍬形蟲 Lucanus datunensis 是臺灣島內體型最小且最受威脅的特有深山鍬形蟲, 只出現在陽明山國家公園大屯山區的高草原區. 親緣分析指出大屯姬深山鍬形蟲和大陸的深山鍬形蟲 L. fortunei 是最近的姐妹種, 粒線體基因多樣性低顯示大屯姬深山鍬形蟲的有效族群量低. (Lin et al 2009 ).
大屯姬深山鍬形蟲 (彭瑄玉繪).
4. 特徵演化
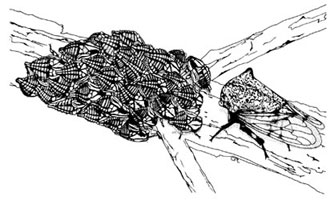
Metcalfiella nigrihumera 角蟬雌蟲守護四到五齡群聚在樹幹上的幼蟲 (drawing by CPL).
許多角蟬種類是群居性的, 角蟬的社會行為變異可以從獨居型,幼蟲和成蟲聚居型,到有高度雌蟲護幼行為 (亞社會行為) 的物種. 角蟬的亞社會行為 (subsociality) 侷限於雌蟲照顧卵或幼蟲, 其中最常見的是雌蟲護卵行為. 角蟬亞科中相近物種社會行為的變異提供了一個探討昆蟲社會行為演化很好的課題 (Lin et al 2004, Lin 2006). 研究成果中角蟬護幼行為的比較分析被引用於一本經典的動物行為學教科書中 (Chap. 12, p. 406-08, Animal Behavior: An Evolutionary Approach, 8th ed. 2005 by John Alcock)

Guayaquila 雌蟲守衛產於樹枝中且用臘質分泌物覆蓋的卵, Photo by CPL.
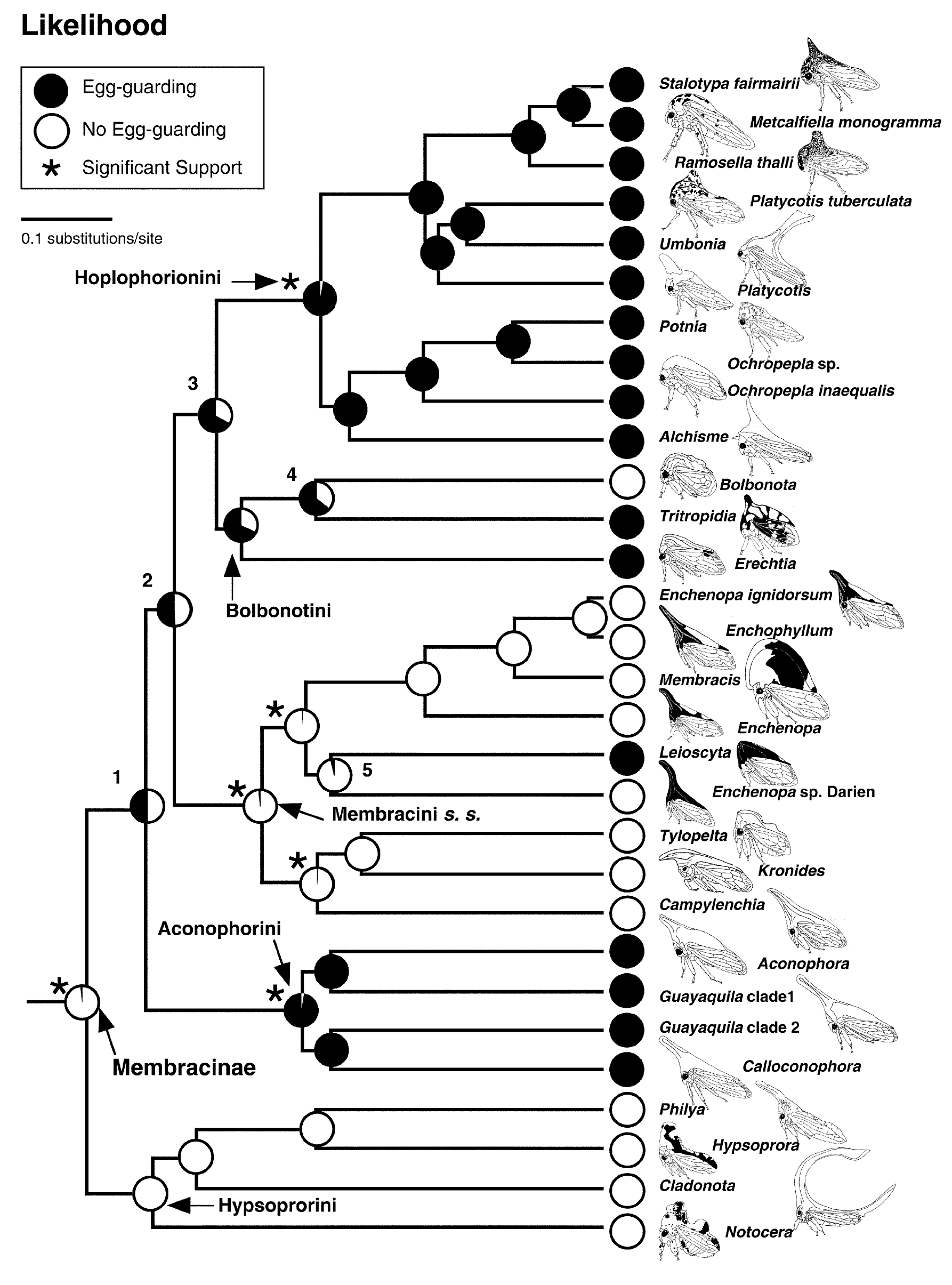
角蟬護幼行為的演化模式 (Lin et al 2004).
5. 親緣地理
i. 北美洲橡樹角蟬

2001 年在宏都拉斯 La Union 拍攝 Platycotis tuberculata 角蟬的未成熟雄蟲 (左) 和雌蟲 (右), Photo by CPL.
透過研究自然族群中基因親緣樹現存的地理分佈, 親緣地理學提供了一個整合性概念架構來探討物種形成和分化的歷史過程. 我們研究北美洲橡樹角蟬 Platycotis vittata 的親緣地理, 並測試其遺傳結構是否符合物種時間隔離的假說 (Lin et al 2007). 親緣地理分析顯示北美橡樹角蟬族群內的遺傳變異顯著, 此遺傳結構與美國東部, 歐札克台地, 和美國西部親緣類群大尺度的地理分佈吻合, 美國東部類群內的北邊和南邊族群並無明顯的親緣地理結構和基因交流, 這個東部族群內無固定遺傳變異的結果符合時間隔離假說的預期.
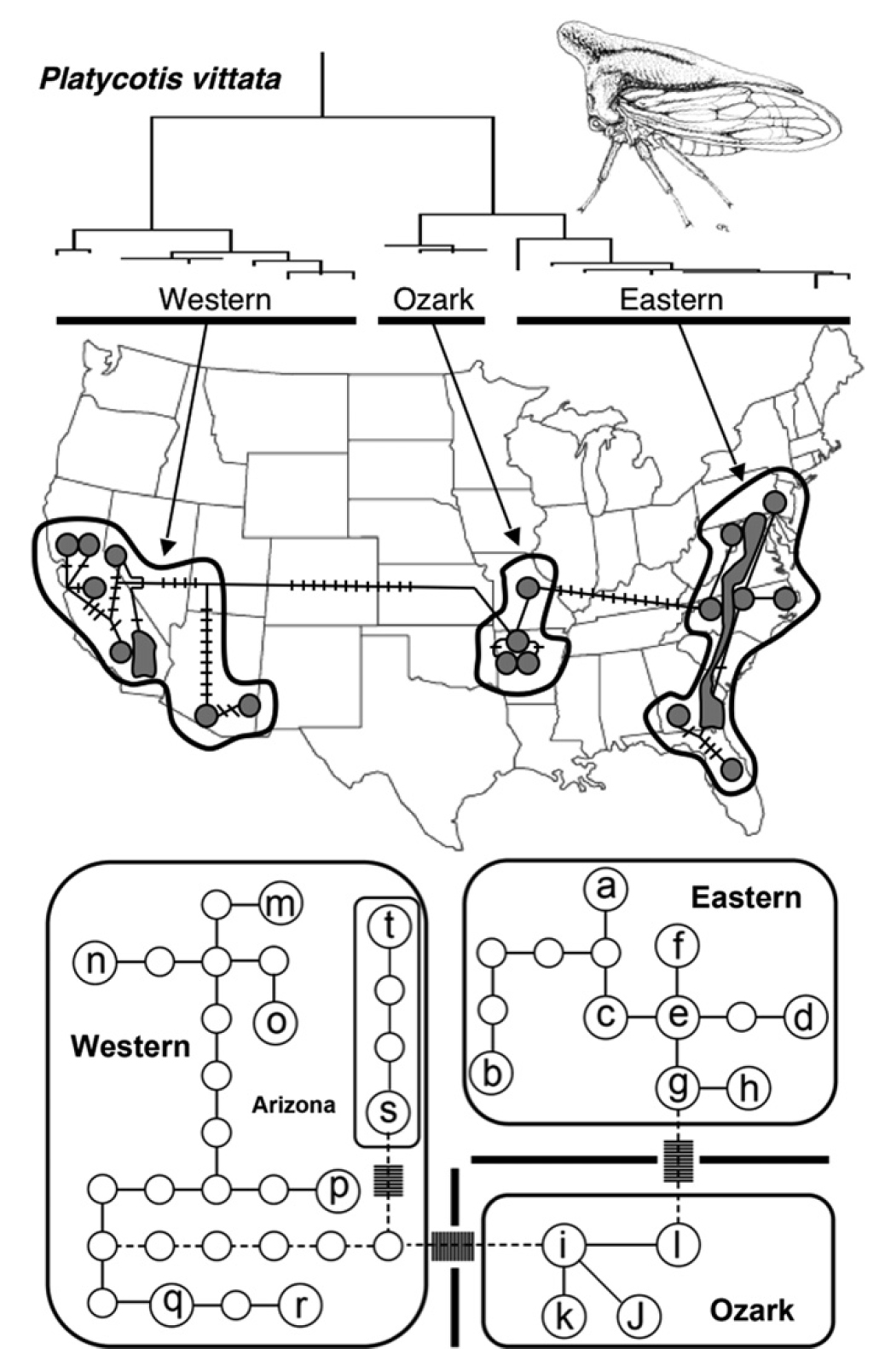
北美洲橡樹角蟬的親緣地理結構 (Lin et al 2007).
ii. 菲律賓群島
研究撰寫中...
iii. 台灣特有昆蟲
台灣在所有亞熱帶地區中地位特殊, 因為她是緊臨熱帶地區南北緯23度唯一具規模的島嶼. 我們研究台灣島上特有種昆蟲的親緣地理來了解這些物種的起源和多樣化.
a. 臺灣深山鍬形蟲 Lucanus formosanus 親緣地理與大顎演化

臺灣深山鍬形蟲 (彭瑄玉繪).
臺灣深山鍬形蟲起源於 1.6 百萬年前更新世初期, 包含阿里山和廣泛分布的兩個在地理上重疊的系群. 大約 20 萬年前, 廣泛分佈的這個系群可能因溫帶森林海拔分布的轉變而產生劇烈地族群擴張. 中央山脈周圍的族群間發生環狀的基因交流, 顯示出這些山脈對臺灣深山鍬形蟲族群形成一個阻斷東西向基因交流的地理障礙. 臺灣深山鍬形蟲族群間的大顎變異程度和預期中的中性分子演化不一致, 顯示性擇或天擇最有可能朔造這些作為武器的大顎在形態上的分化與演化 (Huanag & Lin 2010).
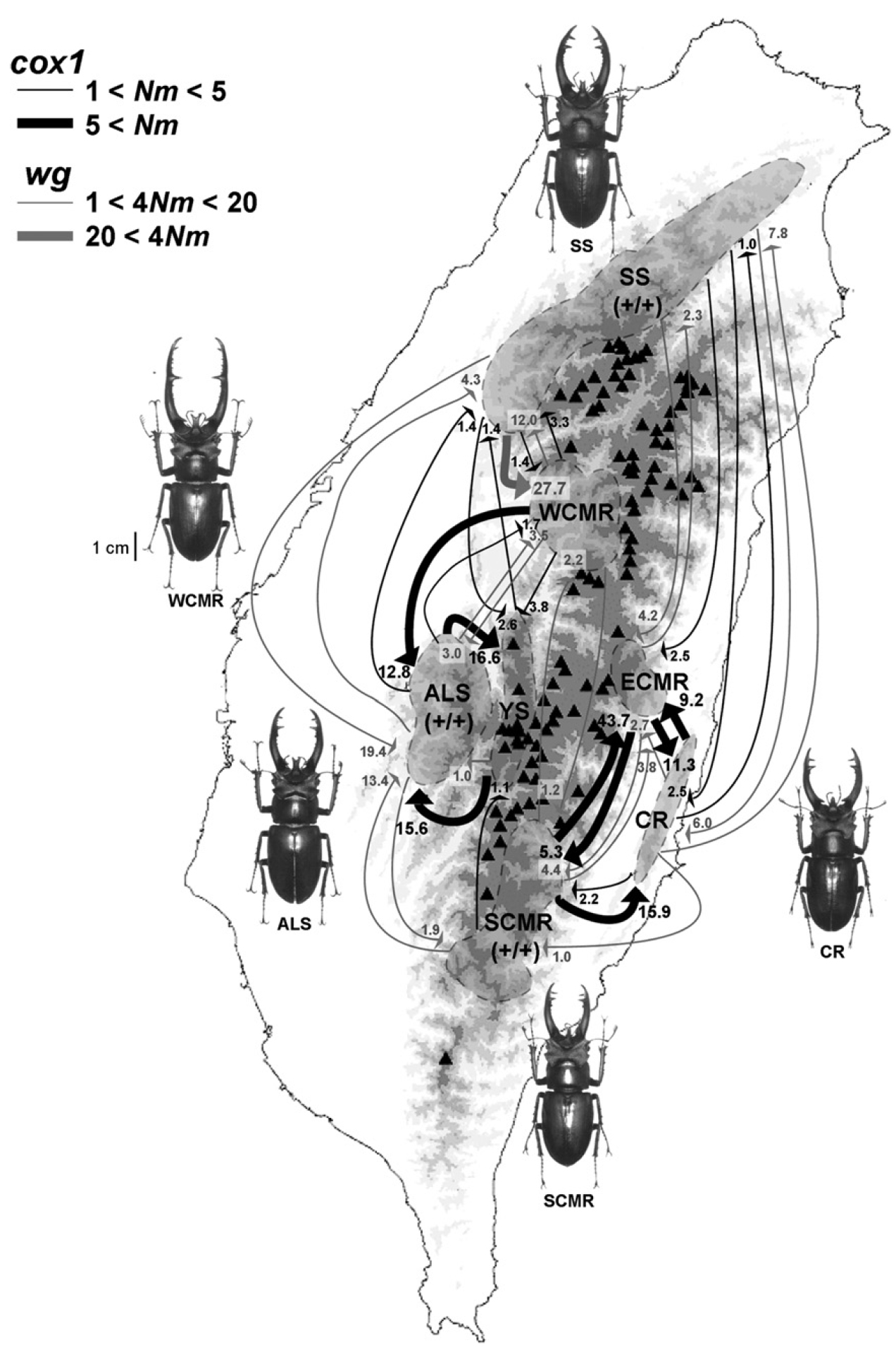
臺灣深山鍬形蟲的基因交流 (Huanag & Lin 2010).